ENVIRONMENTAL VARIATION OF THE POPTELLA COMPRESSA DIET, A XINGU RIVER CHARACIDAE, BRAZILIAN AMAZON
REGISTRO DOI: 10.69849/revistaft/th102412301506
Wilson Martins da Silva1
Janice Muriel Fernandes Lima da Cunha2
RESUMO
Este estudo teve como objetivo caracterizar o efeito da variação ambiental sobre os hábitos alimentares de Poptella compressa nos períodos de seca e cheia no Igarapé Trindade, afluente da margem esquerda do Rio Xingu, em Altamira, Pará. As coletas dos exemplares foram realizadas em uma localidade situada próxima à Rodovia Transamazônica, no mês de fevereiro e setembro de 2010, antes da implantação da barragem de Belo Monte. A metodologia usada constou de captura dos exemplares com tarrafa e rede de emalhar, fixação dos indivíduos em formalina e conservação em álcool para análise posterior em laboratório. A dieta da espécie foi determinada através da análise do conteúdo estomacal, utilizando-se os métodos de freqüência de ocorrência e volume, sendo expressa pelo Índice Alimentar (IAi). Os resultados observados mostraram que o item mais importante na alimentação se constitui de material vegetal no período de estiagem e insetos no período chuvoso. Inferiu-se a alteração da dieta de acordo com a mudança ambiental, sendo caracterizada como onívora tendendo a herbivoria na estiagem e onívora tendendo a insetivoria no período chuvoso.
Palavras-chave: Dieta. Poptella. Variação Ambiental. Ictiofauna.
ABSTRACT
This study aimed to characterize the effect of environmental variation on the feeding habits of Poptella compressa during the dry and wet seasons in the Trindade Stream, a tributary of the left bank of the Xingu river, in Altamira, Pará. The specimens were collected in a location close to the Trans-Amazonian Highway, in February and September 2010, before the construction of the Belo Monte dam. The methodology used consisted of capturing the specimens with a cast net and gillnet, fixing the individuals in formalin and preserving them in alcohol for later analysis in the laboratory. The diet of the species was determined by analyzing the stomach contents, using the frequency of occurrence and volume methods, and expressed by the Food Index (AI). The results showed that the most important item in the diet consists of plant material during the dry season and insects during the rainy season. It was inferred that the diet changed according to environmental changes, being characterized as omnivorous tending towards herbivory in the dry season and omnivorous tending towards insectivory in the rainy season.
Keywords: Diet. Poptella. Environmental Variation. Ichthyofauna.
1. INTRODUÇÃO
Poptella é um gênero da da família Characidae, subfamília Stethaprioninae, da classe Actinopterygii, conhecido como “piaba, piaba-matupiri”, na região do rio Xingu. Está distribuído por várias bacias sulamericanas como Paraná, São Francisco, Tocantins, Amazonas e Orinoco.
Atualmente são consideradas quatro espécies válidas de Poptella: Poptella compressa; Poptella longipinnis; Poptella paraguayensis e Poptella brevispinna (REIS, 1989). A subfamília Stethaprioninae forma um grupo de peixes pequenos, que possuem umcorpo achatado lateralmente, com um espinho que fica logo à frente da sua nadadeira dorsal. O gênero Poptella possui uma distribuição mais abrangente na subfamília (REIS, 1989).
O gênero Poptella tem demonstrado em estudos realizados em outras bacias um nível de plasticidade trófica muito evidente (RESENDE, 2000; ANJOS, 2005; SANTOS et al., 2009). Plasticidade trófica ou alimentar é o termo usado para explicar a habilidade dos animais em utilizar diversos recursos alimentares disponíveis no meio ambiente. Refere-se a uma característica da ictiofauna neotropical sendo importante em locais onde a disponibilidade é bastante diversificada (PIRES; SOUZA; SILVA, 2024). Contudo, como há poucos estudos sobre essa característica para Poptella compressa na Amazônia, onde a variação dos fatores ambientais é muito marcada pela sazonalidade, alternando de períodos de alta e baixa pluviosidade segundo Neto et al. (2021).
Zuanon (1999), em estudos na Volta Grande do rio Xingu, analisou a dieta de três grupos de peixes que ocorrem em cachoeira e encontrou um comportamento trófico influenciado pela sazonalidade, o que ressalta a importância da ecologia trófica para o entendimento do ecossistema aquático desta bacia, que está sujeita a alterações importantes dos fatores ambientais, onde o fator pluviosidade é um dos principais (NETO et al., 2021).
A partir do conhecimento da dieta dos peixes de uma comunidade e da abundância específica dos itens consumidos as diferentes categorias tróficas podem ser identificadas, e o grau de importância dos diferentes níveis tróficos no funcionamento de um determinado sistema, sendo possível entender melhor algumas inter-relações entre os componentes da comunidade (ZAVALA-CAMIN, 1996). A maioria dos peixes pode mudar de um alimento para outro tão logo ocorram alterações na abundância relativa do recurso alimentar em uso segundo Caldatto; Dias; Ferreira (2023), Brazil‐Sousa et al. (2024), Pires; Souza; Silva (2024) e Sena; Súarez (2024).
O estudo da dieta através da análise do conteúdo estomacal apresenta muitos problemas, entre eles o reconhecimento dos organismos que foram ingeridos e sua quantificação (HYNES, 1950). Desta forma, o estudo sobre os hábitos alimentares da ictiofauna permite conhecer exatamente qual o recurso está sendo utilizado pelas diferentes espécies de peixes e, portanto, a importância desses recursos para a manutenção da dinâmica trófica que possibilita a existência da comunidade (MANNA; REZENDE, 2021). Nesse sentido, o objetivo desse estudo foi verificar o efeito da variação ambiental do ciclo hidrológico na ecologia trófica desse grupo no local contribuindo para uma avaliação do estado atual da comunidade íctica, uma vez que estes ambientes são muito vulneráveis aos impactos antrópicos.
2. MATERIAL E MÉTODOS
O estudo foi realizado no igarapé Trindade, também conhecido como Açaizal, que é um afluente da margem esquerda do Rio Xingu. Foram realizadas quatro coletas, duas nos meses de fevereiro (cheia) e duas em setembro (seca) no ano de 2010. Foram capturados 71 exemplares no ponto de coleta situado próximo à BR-230, Rodovia Transamazônica (S: 3°10’21″ e O: 52°08’37″), em zona de remanso do igarapé (Figura 1). Os apetrechos usados para a captura dos exemplares foi tarrafa de malha 12 mm e rede de emalhar com 12 mm entre nós, em aproximadamente uma hora de pesca. Após a amostragem, cada indivíduo foi fixado em formol 10% e depois preservado em álcool 70% para análise posterior em laboratório. Os exemplares testemunhos estão depositados no laboratório de Biologia da Universidade do Estado do Pará-UEPA, Altamira, Pará.

Figura 1 – Local do ponto de coleta no Igarapé Trindade
Fonte: Googlemaps.com
2.1 Identificação da área de estudo
O igarapé Trindade possui tamanho aproximado de 15 km de extensão, profundidade média no local de coleta de 1,5 na seca e 2,3 na cheia. A vegetação marginal é típica de floresta secundária com desenvolvimento clímax bem acentuado. A água apresenta cor transparente durante a seca e mais turva na cheia, com temperatura média de 26° C, PH de 5,9 e oxigênio dissolvido com 49% 02.
Na área de coleta, o dossel apresenta-se mais aberto, proporcionando boa luminosidade ao leito do igarapé, evidenciando o crescimento de algas filamentosas como Spyrogira sp.,(Chlorophyta). Estas algas ocorrem geralmente próximo às margens do riacho e sobre as folhas de macrófitas como um recurso alimentar para peixes herbívoros. Cyperaceae semi-aquática, enraizada no fundo, com folhas e inflorescências aéreas, faz-se presente nas margens do igarapé. No substrato de fundo, geralmente muitos galhos presentes, sendo refúgio para indivíduos da ictiofauna no local, como Leporinus friderici, popularmente Piau-cabeça-gorda e Pterophyllum scalare, conhecido como Acará-bandeira. No fundo do riacho, também se caracteriza por um substrato arenoso com cascalhos e algumas pedras separadas.
2.2 A Ictiofauna do local
No ponto de coleta, foram capturadas um total de 11 espécies, fazendo parte de cinco famílias, sendo 10 da ordem Characiformes (famílias Characidae, Curimatidae, Acestrorhynchidae e Anostomidae) e uma da ordem Perciformes (família Cichlidae), totalizando 123 espécimes.
2.3 Sexo, peso e comprimento padrão
Os indivíduos coletados apresentaram um equilíbrio no número de machos e fêmeas na seca e um número maior de machos na cheia, sendo que não foi observado dimorfismo sexual na espécie. Os sexos foram diferenciados pela análise das gônadas. Os indivíduos analisados também apresentaram peso médio de 4,84 gramas na seca e 10,66 gramas na cheia. O comprimento padrão médio observado foi de 5,74 na seca e 7,85 na cheia o que mostra um aumento de peso e comprimento no mês de fevereiro (tabela 1 e 2).
Tabela 1 – Sexo dos exemplares de P. compressa
Sexo Seca Cheia Total Fi% Machos 18 20 38 61,29 Fêmeas 14 10 24 38,7
Fonte: Autor (2010)
Tabela 2 – Peso e Comprimento Padrão (média geral)
Período Peso Médio (g) Comp. Padrão (cm) Seca (32 indivíduos) 4,84 5,74 Cheia (30 indivíduos) 10,66 7,85
Fonte: Autor (2010)
2.4 Análise de conteúdo estomacal e protocolos utilizados
Neste estudo, após a coleta, os indivíduos foram anestesiados com Eugenol, diluído antecipadamente em etanol e feito um ajuste para uma concentração resultante de 40 mg L-1 (DUARTE et al. 2022), fixou-se cada exemplar em formol 10% e depois conservado em álcool 70% para posterior estudo. Depois, os indivícuos tiveram seus estômagos removidos por incisão látero-ventral para análise em laboratório. Procedeu-se a análise de 62 estômagos, com grau de repleção três (cheios), sendo descartados 9 estômagos que se encontravam vazios, portanto com grau de repleção igual a zero. Para obtenção dos dados qualitativos, o conteúdo estomacal foi analisado sob microscópio estereoscópico e os itens alimentares identificados até o nível taxonômico mais refinado possível. Utilizou-se para essa identificação uso de bibliografia especializada e consulta aos especialistas. Os volumes foram obtidos pela compressão do material analisado com lâmina de vidro sobre placa milimetrada até uma altura de 1mm (HELLAWELL; ABEL, 1971) e o resultado convertido em mililitros (1mm³ = 0,001ml). Em seguida, foi estimado o percentual de cada item em relação ao volume total.Desse modo, foram caracterizados os itens preferenciais para alimentação de P. compressa, segundo o Método de Ocorrência (Fi) e Método Volumétrico (Vi), (HYSLOP, 1980), combinados ao Índice de Importância Alimentar (IAi), (KAWAKAMI; VAZZOLER, 1980), descrito pela equação:
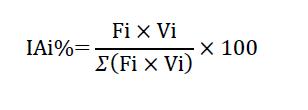
Onde, i = 1, 2… n= determinado item alimentar;
Fi= Freqüência de ocorrência do item alimentar;
Vi= Volume do item alimentar.
Este índice alimentar nos permite diferenciar de forma mais adequada a importância relativa de cada item, qualquer que seja sua condição quanto à frequência de ocorrência e de volume (KAWAKAMI; VAZZOLER, 1980).
Os resultados dos índices alimentares observados demonstraram que P. compressa possui um comportamento trófico que varia de acordo com as alterações dos ciclos hidrológicos.
3. RESULTADOS
Observou-se, neste estudo, uma grande diversidade de itens alimentares consumidos por P. compressa. Pois, como a espécie apresenta plasticidade trófica, variando sempre sua dieta em função da variação ambiental nos períodos de seca e cheia, sempre poderá haver o acréscimo de itens não ainda identificados para o grupo.
3.1 Hábitos alimentares durante a cheia
No período das coletas realizadas no mês de fevereiro, mostrou-se um elevado nível de precipitação que influenciou o nível dos rios na região do município de Altamira, como mostram os dados em Franco et al. (2015), sendo que a fluviometria ficou próximo de 600 cm (Figura 2). Nesse período, P. compressa apresentou uma dieta onívora com forte tendência à insetivoria como mostra a Tabela 3. Os insetos (IAi = 65,97%) representam o alimento mais importante da espécie no período de cheia. O material vegetal (IAi = 15,5%) e as algas filamentosas (IAi = 14,45%) aparecem em segundo e terceiro lugar respectivamente. Os insetos alóctones representam a grande maioria desse item, sendo constituídos de principalmente de Hymenoptera (Formicidae). Foi observada também a presença de um percentual de insetos autóctones representados principalmente por larvas de Ephemeroptera, Chironomidae e Trichoptera.
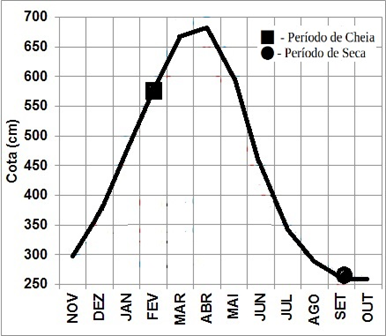
Figura 2 – Evolução da cota média mensal do nível fluviométrico (cm) do rio Xingu no período 1979-2012.
Fonte: Adaptado de Franco et al. (2015)
Tabela 3 – Dieta de P. compressa na Cheia
Item Alimentar Fi% Vi% IAi% Fragmento Vegetal 15,5 28,5 15,5 Algas Filamentosas 8,7 2,2 14,45 Insetos 29,1 47,4 65,97 Outros Invertebrados 4,9 5,1 1,19 Escamas 20,4 14,8 0,92 Detritos/Sedimentos 21,4 1,9 1,97 Total 100 100 100
Fonte: Autor (2010)
3.2 Hábitos alimentares durante a seca
No mês de setembro, observou-se um período de pouca chuva de acordo com Franco et al. (2015), sendo que a fluviometria variou por volta de 250 cm (Figura 2), nesse período. Nesses meses de seca, P. compressa apresentou uma dieta onívora com tendência a herbivoria como mostra a Tabela 4. Material vegetal (IAi = 59,3%) aparece como o item alimentar mais importante da espécie nesse período. O material vegetal observado é composto de muitos pequenos fragmentos de caule, raiz, sementes e folhas de macrófitas. Algas filamentosas (IAi = 13,9%), encontradas na dieta, fazem parte, em sua grande maioria, de Spirogyra sp. e outras da divisão Chlorophyta, além da ocorrência do grupo Rhodophyta. Os insetos (IAi = 18,6%), aparecem como segunda fonte alimentar nesse período. Os insetos alóctones representam a grande maioria desse item, sendo constituídos principalmente de Hymenoptera (Formicidae). Foi observada também uma ocorrência importante de insetos aquáticos representados principalmente por larvas de Ephemeroptera, Chironomidae e Trichoptera.
Observa-se também a presença constante, mas de pouca importância alimentar, de escamas (IAi = 1,8%), o que revela o hábito lepidófago de P. compressa. Portanto, a ocorrência de escamas foi observada, principalmente no período da seca, onde P. compressa possui um menor desenvolvimento quanto ao peso e ao tamanho. A contribuição de outros invertebrados (IAi = 4,3%) mostra que ocasionalmente a espécie pode incluir, em sua dieta, outros itens disponíveis no habitat. Foram encontrados itens ocasionais na dieta como os ácaros da sub-ordem Hydracarina e os poríferos da classe Demospongiae. Essas esponjas que fazem parte da sub-ordem Spongillina, são típicas de água doce e apesar do registro de apenas uma ocorrência destes invertebrados.
Tabela 4 – Dieta de P. compressa na Seca
Item Alimentar Fi% Vi% IAi% Fragmento Vegetal 22 47 59,3 Algas Filamentosas 23,7 10,1 13,9 Insetos 20,3 16 18,6 Outros Invertebrados 3,4 22,5 4,3 Escamas 13,6 2,3 1,8 Detritos/Sedimentos 16,9 2,1 2 Total 100 100 100
Fonte: Autor (2010)
4. DISCUSSÃO
No período de cheia, P. compressa se mostra uma espécie oportunista que se alimenta principalmente dos insetos que caem da vegetação marginal ou são carreados pela chuva e posteriormente levados pela correnteza, sendo formados principalmente por Formicidae. Contudo, neste período, também sua dieta contém uma parcela de insetos autóctones formados principalmente por larvas de Ephemeroptera, Trichoptera e Chironomidae. Destes grupos, é importante destacar a presença de larvas de Trichoptera e Ephemeroptera, pois são insetos utilizados como bioindicadores ambientais importantes para se avaliar as condições da qualidade da água (CALLISTO et al., 2001).
A presença de insetos aquáticos é um aspecto que leva a conclusão de que, no momento, as condições ambientais ainda são toleráveis no local, uma vez que esses invertebrados são muito sensíveis a ambientes poluídos (BERNARDI et al. 2022). Material vegetal e algas filamentosas possuem um índice de importância menor na cheia. Isso se deve a alguns fatores como o aumento da pluviosidade que eleva a nível do igarapé e isso o torna mais caudaloso. As espécies adaptadas aos ambientes aquáticos de remanso fogem para locais onde a velocidade da água é menor. Apesar disso, posicionam-se como fontes alimentares importantes, mas em menor grau em relação aos insetos.
A ocorrência de detritos/sedimentos no estômago de P. compressa pode ser interpretada pela frequencia de insetos aquáticos, seria esperado notar a ocorrência de fragmentos de areia, fragmentos vegetais, pois, sabe-se que Trichoptera usa um casulo construído com esses materiais na fase de larva, localizando-se no fundo do igarapé, é um recurso alimentar, de acordo com Garcia; Lopez; Ibagon (2023).
Observou-se que P. compressa é uma espécie que se desloca em cardumes constituídos em torno de 15 a 20 indivíduos. Isso está de acordo com Cassati (2002) ao ressaltar que, geralmente, Poptella compressa, forma cardumes de 10 a 25 indivíduos que nadam ativamente à meia-água coletando partículas arrastadas pela corrente. Durante a noite, os indivíduos estão parados nas partes mais profundas dos igarapés, mas podem em algumas ocasiões se alimentar de insetos que caem na água, provenientes da mata ciliar (LEITE et al., 2023).
Considerando-se o aspecto da sazonalidade, as mudanças no ciclo hidrológico, determinam variações nos itens alimentares utilizados por diferentes espécies em função de períodos de chuva e seca (ESTEVES; ARANHA; ALBRECHT, 2021). Este estudo corrobora os dados já publicados sobre alterações nas dietas de peixes em face às mudanças ambientais, confirmando a consistente plasticidade alimentar de Poptella (SENA; SÚAREZ, 2024)
Com relação ao perfil trófico da espécie, Santos et al. (2009), inferiu para Poptella paraguayensis uma dieta onívora com uma forte tendência a herbivoria, porém encontraram nesse mesmo estudo um baixo percentual de insetos. No entanto, esses resultados foram baseados na análise de apenas nove estômagos sem a abordagem da sazonalidade que pode determinar a plasticiade trófica na comunidade da ictiofauna de acordo com Caldatto; Dias; Ferreira (2023) e Pires; Souza; Silva (2024).
Trabalhando em áreas distintas, Resende (2000), considerou essa mesma espécie como zooplanctófoga/insetívora. Os resultados do presente estudo mostram que P. compressa apresenta no local analisado plasticidade trófica. No período de seca o alimento mais importante para a espécie foi material vegetal e algas filamentosas, com insetos em segundo lugar, portanto uma dieta onívora com tendência a herbivoria. No entanto, na cheia, a dieta onívora que tende à insetivoria o que mostra de forma consistente que ela pode mudar sua alimentação em função dos recursos disponíveis de acordo com as mudanças ambientais (BEGON; TOWNSEND, 2023).
Segundo Anjos (2005), P. compressa apresenta uma dieta insetívora para a espécie. Contudo, a análise dos hábitos alimentares foi baseada no conteúdo estomacal de apenas sete indivíduos e também o elemento sazonal não foi avaliado. O presente estudo, baseado em um número bem maior de estômagos analisados, corrobora a insetivoria dentro de um perfil onívoro para espécie, mas apenas no período de cheia. Na seca, tende a herbivoria.
A presença de esponja na dieta de P. compressa é relevante, apesar do registro único, pois há pouco conhecimento sobre esse grupo na Amazônia. Portanto, sugere-se que um estudo sobre taxonomia e sistemática seja relevante através da coleta de conteúdo estomacal de peixes, usando a metodologia proposta por Volkmer-Ribeiro (1981), pela análise do conteúdo intestinal de anostomídeos, que são conhecidos comedores de poríferos na região do Xingu (ZUANON, 1999).
As espécies mudam sua dieta em ambientes onde as alterações ambientais são marcantes. Isso foi verificado por vários estudos como Caldatto; Dias; Ferreira (2023), Brazil‐Sousa et al. (2024), Pires; Souza; Silva (2024) e Sena; Súarez (2024), onde pode-se observar o papel da variação ambiental estabelecida na alteração do ciclo hidrológico tendo um efeito direto sobre a dieta da ictiofauna.
5. CONCLUSÃO
Pode-se inferir no presente estudo que P. compressa apresenta no local analisado plasticidade trófica que muda sua alimentação em função das alterações ambientais do ciclo hidrológico. No período de seca o alimento mais importante para a espécie foi material vegetal e algas filamentosas, com insetos em segundo lugar, portanto uma dieta onívora com tendência a herbivoria. No entanto, na cheia, a dieta onívora que tende à insetivoria o que mostra de forma consistente que ela pode mudar sua alimentação em função dos recursos disponíveis de acordo com as mudanças nos ciclos hidrológicos. Finalmente, é possível inferir também que a presença de uma entomofauna aquática demonstra o estado ainda relativamente saudável das águas no trecho estudado.
REFERÊNCIAS
ANJOS, M. B. Estrutura de comunidades de peixes de igarapés de terra firme na Amazônia Central: composição, distribuição e características tróficas. Dissertação de Mestrado, Universidade Federal do Amazonas – Instituto Nacional de Pesquisas da Amazônia (INPA). Manaus, Amazonas. 68pp. 2005.
BEGON, M.; TOWNSEND, C. R. Ecologia: de indivíduos a ecossistemas. Artmed editora, 2023.
BERNARDI, João P.; LIMA-REZENDE, C. A.; SANTOS, J. A.; REZENDE, R. S. Educação ambiental por invertebrados bioindicadores de qualidade de água no Oeste de Santa Catarina. REMEA-Revista Eletrônica do Mestrado em Educação Ambiental, v. 39, n. 1, p. 265-283, 2022.
BRAZIL‐SOUSA, Clarissa; SOARES, B. E.; SVANBÄCK, R.; ALBRECHT, M. P. A especialização individual é maior em populações generalistas de posições tróficas intermediárias a altas em peixes tropicais de água doce. Austral Ecology, v. 49, n. 1, p. e13397, 2024.
CALDATTO, Amanda M.; DIAS, Rosa M.; FERREIRA, Anderson. Diet of Moenkhausia bonita (Benine, Castro & Sabino 2004)(Characiformes: Characidae) in streams in the basin of rio Formoso, Brazilian Midwest. Biota Neotropica, v. 23, n. 2, p. e20221388, 2023.
CALLISTO, Marcos., MORETTI, M., GOULART, M. Macroinvertebrados bentônicos como ferramenta para avaliar a saúde de riachos. Revista Brasileira de Recursos Hídricos, v. 6, n. 1, p. 71-82, 2001.
ESTEVES, K.; ARANHA, J. M. R.; ALBRECHT, M. P. Ecologia trófica de peixes de riacho: uma releitura 20 anos depois. Oecologia Australis, v. 25, n. 2, p. 282-282, 2021.
FRANCO, Vânia S.; Souza, E. B.; Pinheiro, A. N.; Dias, T. S. S.; Azevedo, F. T.; Santos, J. C. C. Evolução mensal da cota fluviométrica do Rio Xingu em Altamira-PA associada aos eventos El Niño e La Niña. Ciência e Natura, v. 37, n. 1, p. 104-109, 2015.
GARCIA, Daniel Andres Feriz; LOPEZ, German; IBAGON, Nicole Estefania. Análisis de contenido estomacal de la sabaleta Brycon henni Eigenmann, 1913 en la parte baja del río Timbío, Cauca-Colombia. Revista Novedades Colombianas, v. 18, n. 1, 2023.
HELLAWELL, J. M.; ABEL, R. A rapid volumetric method for the analysis of the food of fishes. Journal of fish Biology, v. 3, n. 1, p. 29-37, 1971.
HYNES, H. B. N. The food of fresh-water sticklebacks (Gasterosteus aculeatus and Pygosteus pungitius), with a review of methods used in studies of the food of fishes. The journal of animal ecology, p. 36-58, 1950.
HYSLOP, E. J. Stomach contents analysis—a review of methods and their application. Journal of fish biology, v. 17, n. 4, p. 411-429, 1980.
KAWAKAMI, E.; VAZZOLER, G. Método gráfico e estimativa de índice alimentar aplicado no estudo de alimentação de peixes. Boletim do Instituto oceanográfico, v. 29, p. 205-207, 1980.
LEITE, E. F., GODOI, D. S., JACYNTHO, L. A., MACENO, J. F. S., DUARTE, C. R. A. Hábito alimentar e reprodução de Astyanax scabripinnis (Jenyns, 1842) na sub-bacia do Rio Queima-Pé, no município de Tangará da Serra–MT. Brazilian Journal of Development, v. 9, n. 3, p. 10169-10183, 2023.
MANNA, L. R.; REZENDE, C. F. Variação intraespecífica em peixes de riacho com ênfase na ecologia trófica. Oecologia Australis, v. 25, n. 2, p. 322-322, 2021.
NETO, Albertino M.; BATISTA, L. M., SOUSA; M. C., FREITAS, K. M.; ARAÚJO, S. R. Sensoriamento remoto na análise de variáveis ambientais influenciadas pela implantação da usina hidrelétrica de Belo Monte (PA). Caderno Geografia, v. 31, n. 823, p. 2318-2962.2021, 2021.
PIRES, Ingrid C. C.; SOUZA, Fabiane B.; SILVA, André T. Trophic plasticity of tetra-fish, Psalidodon aff. fasciatus (Cuvier, 1819)(Characidae), in an Atlantic Forest stream of Northeast Brazil. Anais da Academia Brasileira de Ciências, v. 96, n. 2, p. e20230652, 2024.
REIS, R. E. Systematic revision of neotropical characid subfamily Stethaprioninae (Pisces, Characiformes). Comun. Mus. Cienc. PUCRS, Ser. zool., v. 2, p. 3-86, 1989.
RESENDE, E. K. Trophic structure of fish assemblages in the lower Miranda river, Pantanal, Mato Grosso do Sul State, Brazil. Revista brasileira de Biologia, v. 60, p. 389-403, 2000.
SANTOS, Claudineia Lizieri; DOS SANTOS, Iracenir Andrade; DA SILVA, Carolina Joana. Ecologia trófica de peixes ocorrentes em bancos de macrófitas aquáticas na baia Caiçara, Pantanal Mato-Grossense. Revista Brasileira de Biociências, v. 7, n. 4, 2009.
SENA, Karoline A.; SÚAREZ, Yzel R. Spatial variation, more than ontogenetic, explains the diet of Bryconamericus exodon in two Pantanal rivers. Acta Limnologica Brasiliensia, v. 36, p. e18, 2024.
VOLKMER-RIBEIRO, C.; GROSSER, K. M. Gut Contents of Leporinus obtusidens “Sensu” von Ihering (Pisces, Characoidei) Used in a Survey for Freshwater Sponges. Revista Brasileira de Biologia, v. 41, n. 1, p. 175-183, 1981.
ZAVALA-CAMIN, Luis Alberto. Introdução aos estudos sobre alimentação natural em peixes. Editora da Universidade Estadual de Maringá, 129p. 1996.
ZUANON, J. História natural da ictiofauna de corredeiras do rio Xingu, na região de Altamira, Pará, In Instituto de Biologia. 199 p. Tese (Doutorado em Ecologia) – Universidade Estadual de Campinas, Campinas, 1999.
1 Docente no curso de Engenharia Ambiental da Universidade do Estado do Pará (UEPA), Campus de Altamira-PA. Mestre em Ecologia Aquática e Pesca pela Universidade Federal do Pará (UFPA). e-mail: wilson.silva@uepa.br
2 Docente no curso de Ciências Biológicas da Universidade Federal do Pará, Campus de Bragança-PA. Doutora em Ciências Biológicas pela Universidade de São Paulo (USP). e-mail: janice@ufpa.br